Maladie de Charcot (SLA) : quand les motoneurones s’éteignent sous le feu croisé du terrain

Introduction
Dans l’univers de la neurologie, la Sclérose Latérale Amyotrophique (SLA) — plus connue sous le nom de maladie de Charcot — est souvent vécue comme un coup de tonnerre. Le diagnostic est abrupt : une paralysie progressive, des muscles qui s'atrophient et un corps qui refuse d'obéir parce que les ordres de mouvement ne passent plus. Face à cela, la médecine conventionnelle se concentre sur le ralentissement des symptômes, sans toujours pouvoir expliquer le pourquoi de cet incendie cellulaire.
Pourtant, si on prend un peu de hauteur grâce aux neurosciences modernes, la SLA s'éclaire d'un jour nouveau. Elle n'est pas un phénomène magique ou isolé. Elle s'inscrit en réalité dans la parfaite continuité des dossiers que nous développons ici :
- après avoir compris comment l'inflammation de bas grade engourdit le cerveau global, ce que l’on appelle le brouillard mental (voir l’article : Inflammation chronique de bas grade : quand le feu reste allumé en permanence),
- après avoir vu comment le système immunitaire peut grignoter l'isolant de nos câbles nerveux dans la sclérose en plaques (voie l’article : Sclérose en plaques (SEP) : Quand l’inflammation et l’épuisement cellulaire perturbent le dialogue du cerveau),
nous braquons aujourd'hui le projecteur sur la cellule nerveuse elle-même.
La SLA est la rupture brutale d'un écosystème. C'est l'histoire de cellules d'élite — les motoneurones — qui finissent par s'éteindre, épuisées par une surchauffe électrique (l'excitotoxicité du glutamate), une panne de carburant généralisée (les mitochondries) et une trahison de leurs propres gardiens (la microglie).
Pour comprendre pourquoi ces cellules sont si vulnérables dans la SLA, il faut d’abord réaliser qu’un motoneurone est une véritable cellule hors norme, soumise à des contraintes biologiques gigantesques.
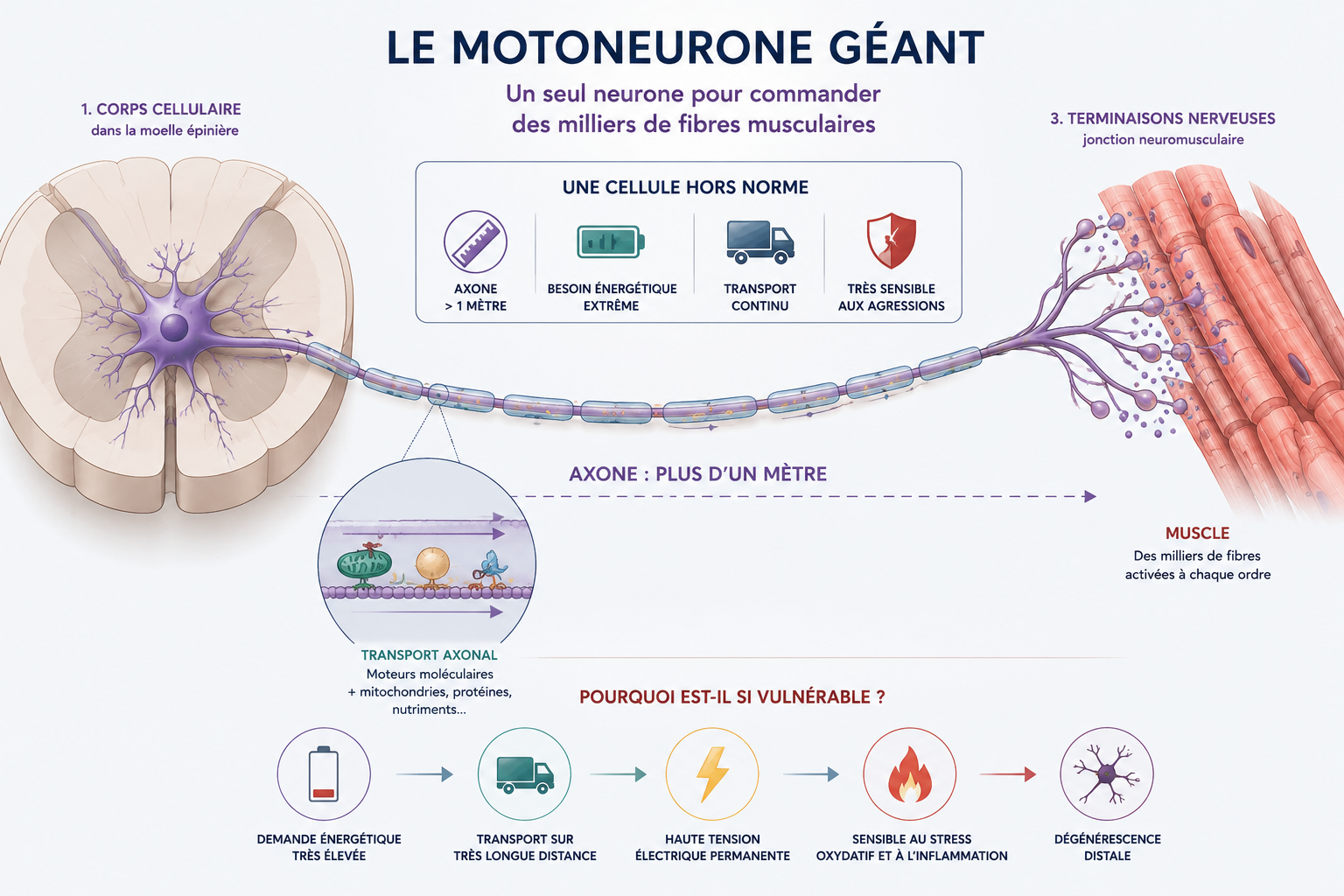
Cette architecture spectaculaire explique pourquoi les motoneurones sont parmi les cellules les plus exigeantes et les plus fragiles du système nerveux : au moindre déséquilibre énergétique ou inflammatoire, toute la chaîne de ravitaillement peut commencer à s’effondrer.
1) Les motoneurones : Des géants cellulaires sous haute tension
Pour comprendre pourquoi la maladie s'en prend spécifiquement aux neurones moteurs (ceux qui commandent nos muscles), il faut imaginer l'incroyable défi physique que ces cellules relèvent chaque seconde.
Dans notre imaginaire, un neurone est une cellule microscopique et ronde. C’est vrai pour notre cerveau. Mais les motoneurones de notre moelle épinière sont des géants.
Pour envoyer un ordre de mouvement du bas de notre dos jusqu'au bout de notre gros orteil, le motoneurone déploie un bras unique et ultra-fin : l'axone. À l'échelle de notre corps, ce bras microscopique mesure parfois plus d'un mètre de long. C’est l’équivalent pour une cellule d’avoir un bras de plusieurs kilomètres !
Cette anatomie hors norme impose deux contraintes biologiques titanesques, qui expliquent leur immense vulnérabilité :
- Une autoroute de ravitaillement saturée (Le flux axonal) : Le cœur du neurone (où sont fabriqués l'énergie, les protéines et les outils de survie) se situe dans la moelle épinière. Il doit acheminer ce matériel jusqu’au bout du pied via de minuscules moteurs moléculaires. Ce flux logistique permanent consomme une quantité astronomique d'ATP (l'énergie cellulaire). Le moindre ralentissement, et l'extrémité du neurone meurt de faim.
- Une ligne à haute tension permanente : Pour propulser un message électrique à plus de 300 km/h au moindre ordre de mouvement, le motoneurone doit maintenir une tension électrique interne phénoménale, jour et nuit.
Par leur structure même, les motoneurones sont donc les formules 1 de notre système nerveux : ultra-performants, mais gourmands, fragiles et dépendants d'un environnement parfait. Si le terrain s'enflamme, ils sont les premiers à basculer.
2) La triade des cercles vicieux : Surchauffe, panne sèche et trahison immunitaire
La mort d'un motoneurone n'est jamais subite. C'est une lente asphyxie provoquée par trois phénomènes qui s'alimentent les uns les autres. Si on ne coupe pas ces boucles, le système s'emballe et la maladie progresse.
2.1) L’excitotoxicité du glutamate : La surchauffe électrique
Pour envoyer un ordre de mouvement aux muscles, le système nerveux utilise un carburant chimique, un neurotransmetteur appelé le glutamate. C'est l'accélérateur du système.
En temps normal, le glutamate donne son impulsion puis est immédiatement nettoyé de la synapse par des cellules d'entretien, les astrocytes, qui possèdent de véritables petites pompes d'aspiration appelées EAAT2.
Dans la SLA, le pied reste bloqué sur l'accélérateur :
- Le système de nettoyage se paralyse : À cause de l'inflammation environnante, les pompes EAAT2 sont paralysées. Le glutamate s'accumule et stagne dans la synapse.
- L'inondation de calcium : Excité en permanence par ce trop-plein de glutamate, le motoneurone panique. Ses portes membranaires s'ouvrent en grand et laissent entrer massivement du calcium. Or, si le calcium est excellent pour les os, il est hautement toxique lorsqu'il s'accumule à l'intérieur d'un neurone. Il agit comme un courant de surtension qui commence à « griller » les composants internes de la cellule.
Dans la SLA, le glutamate agit progressivement comme une pédale d’accélérateur bloquée. Le neurone reste stimulé en permanence, sans parvenir à ralentir correctement son activité électrique.
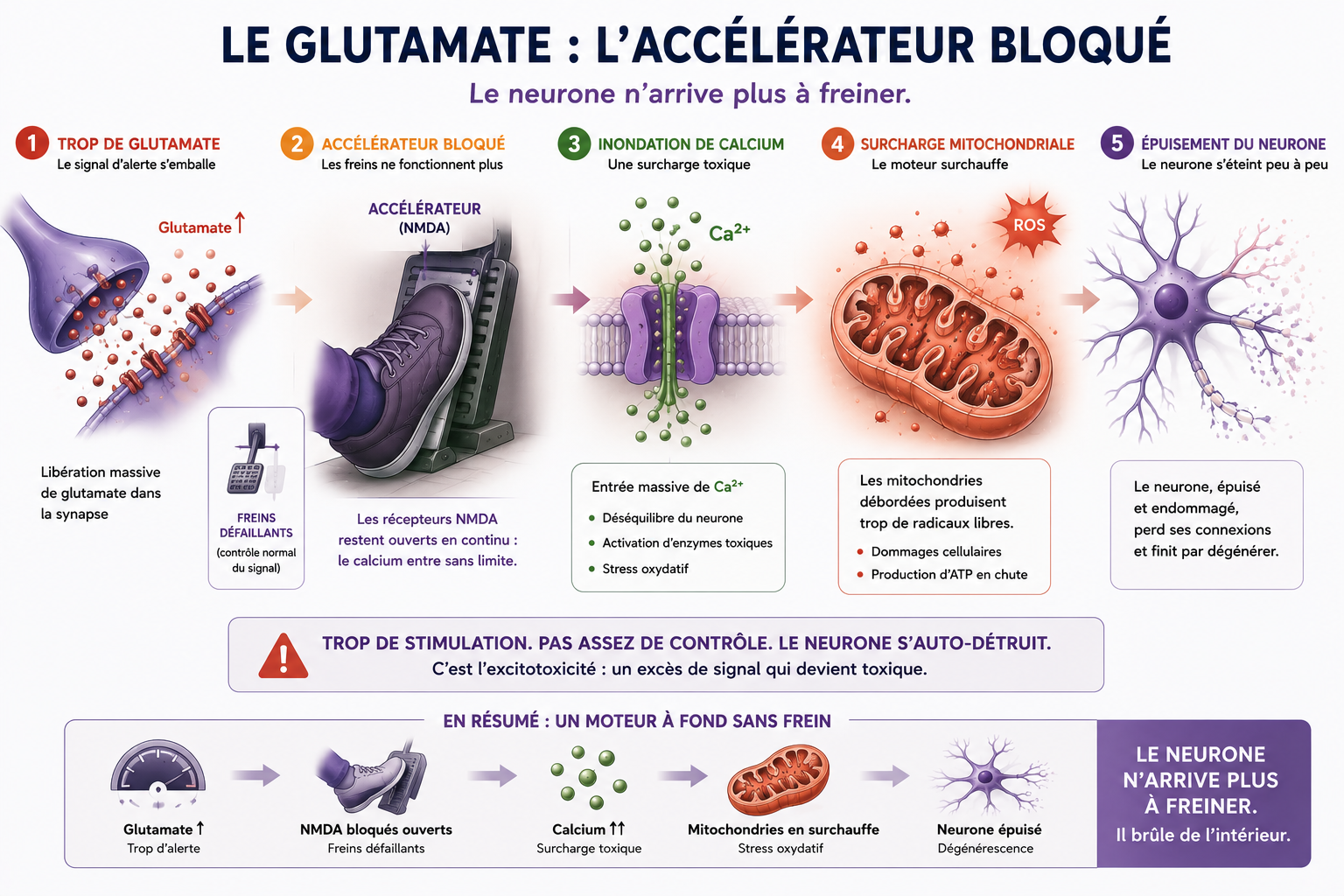
Cette hyperstimulation permanente finit par épuiser les mitochondries, déclencher un stress oxydatif massif et fragiliser progressivement l’ensemble du motoneurone.
2.2) Le naufrage mitochondrial : La panne sèche
Cette agression électrique frappe de plein fouet les usines énergétiques du motoneurone : ses mitochondries. Dépassées par l'inondation de calcium et attaquées par les radicaux libres (ERO), elles capitulent (voir l’article : Fatigue cellulaire : le rôle clé des mitochondries dans notre énergie).
Comme cela a été vu pour la fatigue chronique, la mitochondrie bascule alors dans son mode de survie ancestral : la Réponse de Danger Cellulaire (mode CDR).
Au lieu de fabriquer l'ATP nécessaire pour faire circuler les nutriments le long de son axone d'un mètre, elle coupe les générateurs. Le neurone entre en panne sèche d'énergie. En mourant, la mitochondrie se fissure et rejette des signaux de détresse dans l'espace extracellulaire, des molécules d'alarme appelées DAMPs (comme des fragments de son propre ADN. Voir l'article : Comment notre corps détecte le danger : Les sentinelles invisibles de l’immunité).
2.3) La furie microgliale : Des pompiers qui deviennent pyromanes
C'est ici que le piège se referme au cœur de la moelle épinière (la microgliose spinale). En captant ces DAMPs (les débris de mitochondries en souffrance), les cellules de l'immunité du cerveau (la microglie, vue dans l’article : La microglie : Les gardiens immunitaires du cerveau au cœur de la neuro-inflammation) croient à une agression massive.
Elles basculent instantanément en mode guerrier (le phénotype M1) et se mettent à déverser un torrent de cytokines hautement inflammatoires, comme le TNF-α et l'IL-1β.
2.4) La boucle infernale
Ce torrent de cytokines pro-inflammatoires vient directement bloquer les dernières pompes de nettoyage EAAT2 et modifie l'architecture des récepteurs du neurone (les récepteurs AMPA) pour les rendre encore plus perméables au calcium. L'inflammation microgliale entretient la surchauffe électrique, qui aggrave la panne d'énergie, qui rallume l'inflammation. Le cercle vicieux est bouclé.
À partir de ce moment, plusieurs mécanismes destructeurs commencent à s’auto-alimenter mutuellement. Le glutamate, les mitochondries et la microglie entrent alors dans une véritable triade de cercles vicieux biologiques.
Le motoneurone entre alors dans une spirale biologique auto-entretenue dont il devient extrêmement difficile de sortir.
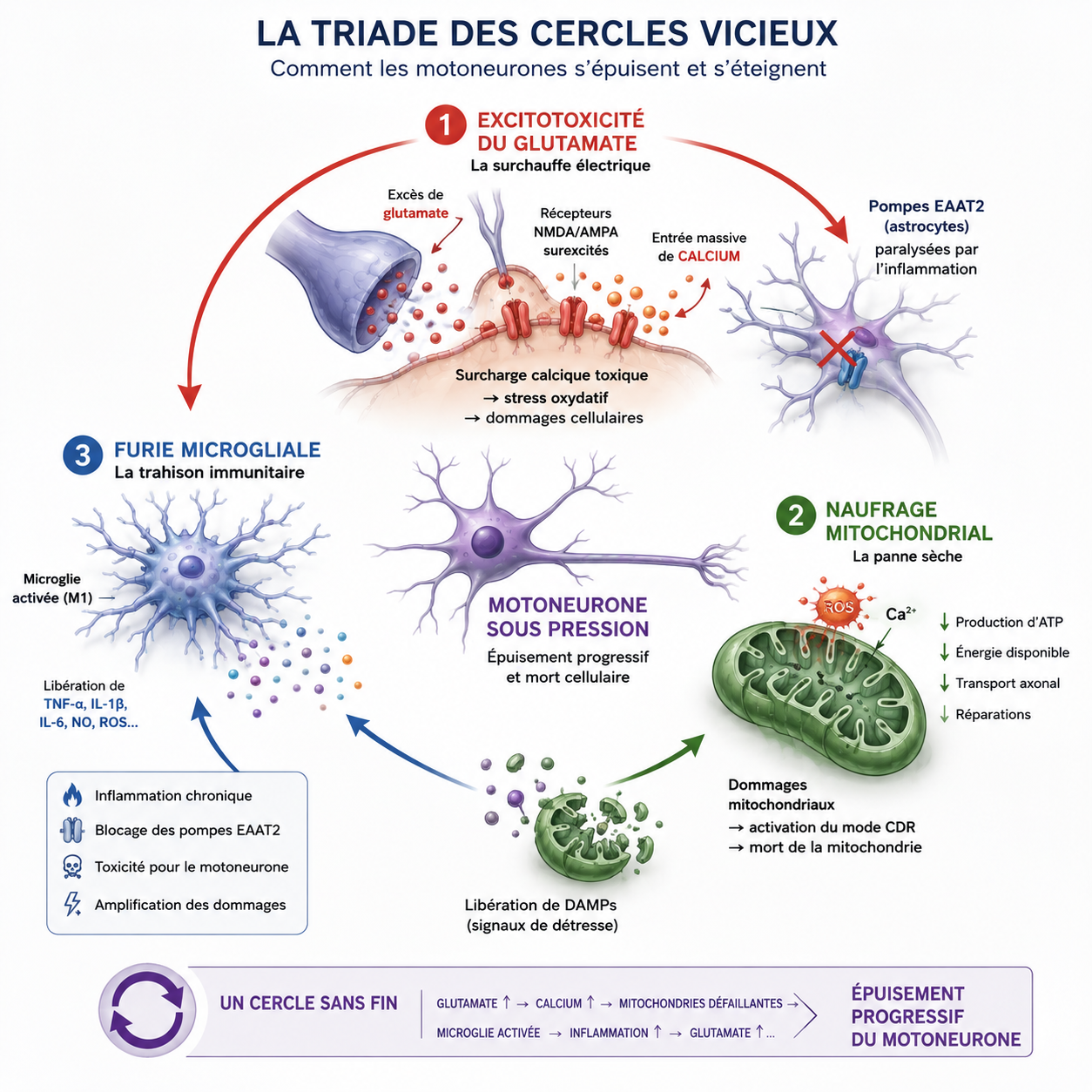
Plus ces boucles s’emballent, plus le motoneurone perd sa capacité à maintenir son équilibre électrique, énergétique et inflammatoire. La cellule finit alors par s’épuiser progressivement jusqu’à la dégénérescence.
Cette défaillance énergétique a une conséquence dramatique : le motoneurone n’est bientôt plus capable d’acheminer correctement l’énergie, les protéines et les outils de réparation le long de son immense axone.
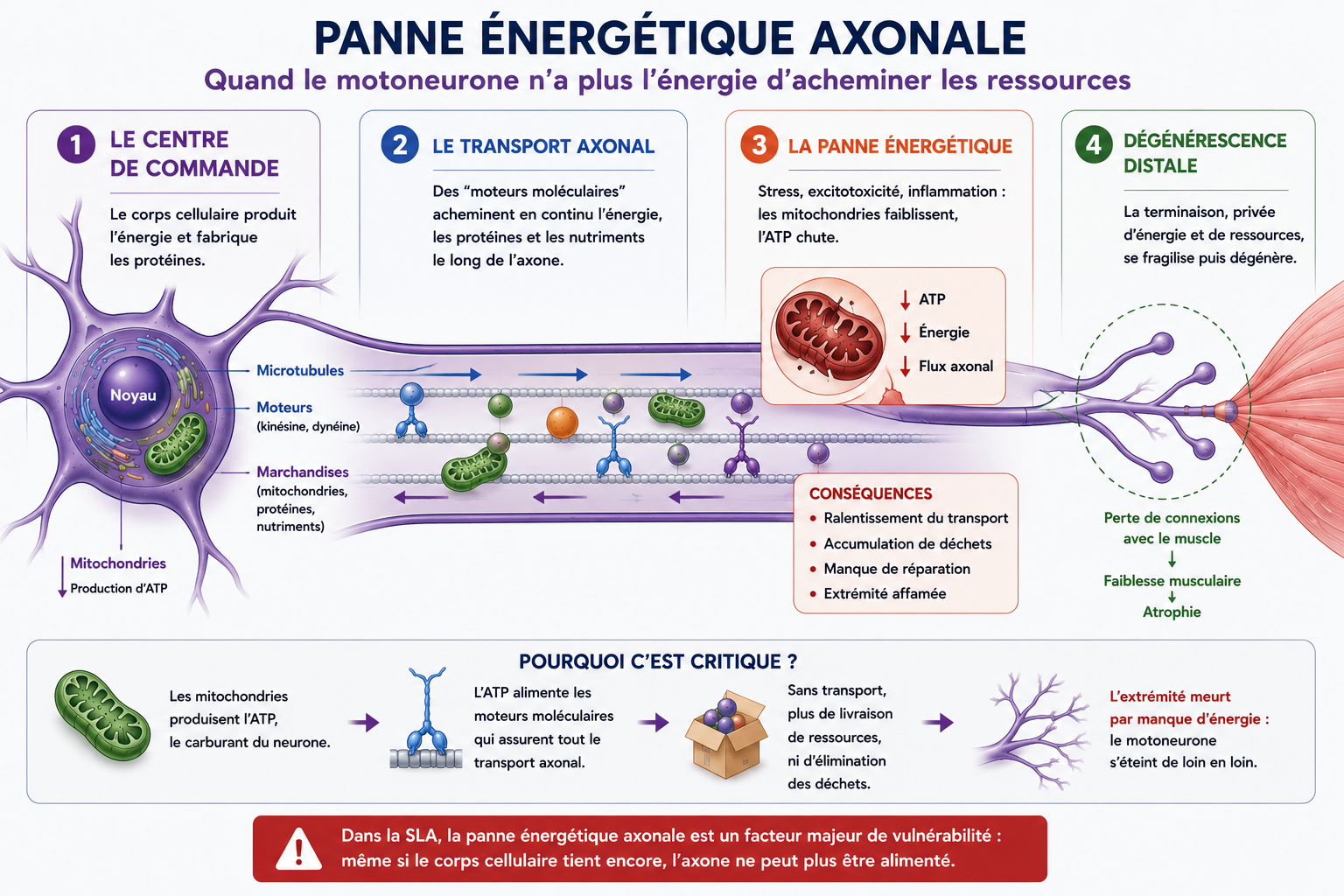
Dans la SLA, le motoneurone ne meurt donc pas uniquement sous l’effet de l’inflammation ou du glutamate. Il s’éteint aussi progressivement par incapacité énergétique à maintenir son propre réseau de transport interne.
3) Génétique vs Épigénétique : Les hypothèses du terrain
Face à l’emballement de ce cercle vicieux, des questions se posent :
- qu’est-ce qui déclenche l'incendie originel ?
- est-ce une fatalité inscrite dans nos gènes, ou un accident de parcours provoqué par notre environnement ?
Les chercheurs estiment aujourd'hui que la SLA est une maladie multifactorielle, où l'inné et l'acquis se croisent de façon complexe.
3.1) La part génétique : Les maillons faibles de l'ADN
La forme purement héréditaire de la SLA reste minoritaire, ne représentant qu'environ 10 % des cas. Dans ces situations, ce sont de petites « fautes d'orthographe » dans notre code génétique qui viennent fragiliser directement la survie du motoneurone. Les scientifiques ont identifié deux coupables majeurs :
- La mutation du gène SOD1 (Le bouclier brisé) : Normalement, ce gène fabrique une enzyme (la Superoxyde Dismutase) qui sert de bouclier pour nettoyer les radicaux libres. Quand ce gène subit une mutation, le bouclier se brise : le motoneurone se retrouve sans défense face à l'oxydation, accélérant le naufrage des mitochondries.
- L'anomalie de la protéine TDP-43 (L'encombrement cellulaire) : Cette protéine est normalement chargée de lire et de trier les messages au cœur du noyau de la cellule. Dans la SLA, elle se dérègle, s'échappe du noyau et s'accumule sous forme de « tas de déchets » toxiques (des agrégats protéiques) dans le cytoplasme du neurone. Cet encombrement finit par étouffer la cellule de l’intérieur.
3.2) La part épigénétique : Quand l'environnement allume la mèche
Pour les 90 % de cas restants — appelés SLA sporadique —, il n'y a pas d'histoire familiale. Le patrimoine génétique de départ est sain, mais ce sont des facteurs environnementaux et des choix de vie (l’épigénétique) qui vont venir modifier l'expression des gènes et saturer les capacités d'adaptation du terrain.
Les chercheurs avancent aujourd'hui plusieurs hypothèses majeures :
- L’exposition aux toxiques environnementaux (Les fausses briques) : L'exposition chronique aux métaux lourds (plomb, mercure) ou à certaines toxines environnementales comme la BMAA (produite par des cyanobactéries présentes dans certaines eaux ou algues) est fortement suspectée. La BMAA a la particularité de ressembler à un acide aminé classique. Le motoneurone l'utilise par erreur comme une brique de construction, ce qui crée des protéines déformées qui finissent par s'agglomérer et saturer la cellule (mimant le problème de la TDP-43).
- L'hypothèse de la "maladie athlétique" (La surchauffe des moteurs d'élite) : C'est un paradoxe qui frappe souvent les esprits : la SLA touche un nombre anormalement élevé de sportifs de haut niveau (footballeurs, cyclistes). L'hypothèse actuelle est qu'une sollicitation physique extrême et chronique pousse les motoneurones à une demande d'énergie permanente. Si ce profil présente par ailleurs une petite fragilité à éliminer les radicaux libres ou une microglie naturellement plus réactive, cette surutilisation électrique prolongée peut faire disjoncter le système et initier la cascade de la maladie.
Ce qu'il faut retenir : La SLA sporadique n'est pas écrite à l'avance. C'est l'accumulation sur un terrain sensible d'une charge toxique, d'un stress oxydatif intense et d'un emballement immunitaire qui finit par forcer les motoneurones à capituler.
4) Soutenir le motoneurone : calmer la surchauffe et protéger l’énergie cellulaire
Face à une pathologie aussi complexe que la SLA, l’aromathérapie clinique ne prétend évidemment pas réparer un motoneurone détruit. En revanche, certaines molécules aromatiques possèdent des propriétés particulièrement cohérentes pour tenter de ralentir l’emballement biologique du terrain (voir l’article : Les actions des Huiles Essentielles (HE) sur le corps humain).
L’objectif n’est donc pas de « guérir » la maladie, mais de soutenir les cellules nerveuses encore fonctionnelles en essayant de :
- calmer la surchauffe électrique liée au glutamate,
- réduire l’incendie microglial,
- protéger les mitochondries,
- et limiter l’effondrement énergétique du neurone.
Dans cette logique, plusieurs familles moléculaires végétales attirent aujourd’hui l’attention par leur cohérence neurobiologique.
4.1) Calmer l’excitotoxicité du glutamate : ralentir la surchauffe du motoneurone
Dans la SLA, le motoneurone se retrouve exposé à une stimulation excessive par le glutamate, le principal neurotransmetteur excitateur du système nerveux. Ce phénomène agit comme une pédale d’accélérateur bloquée : le neurone laisse entrer trop de calcium, s’épuise et finit par basculer en détresse métabolique.
Le linalol contenu dans l’huile essentielle de Coriandre graines possède ici un profil particulièrement intéressant. Cette petite molécule très diffusible semble capable de moduler certains récepteurs du glutamate (notamment NMDA) tout en renforçant l’action apaisante du GABA, le grand frein naturel du cerveau.
Concrètement, le linalol aide à réduire l’hyperexcitabilité neuronale et à limiter l’inondation calcique qui surcharge les mitochondries du motoneurone.
4.2) Désamorcer l’incendie microglial : calmer IL-1β et TNF-α
Lorsque les motoneurones souffrent, ils libèrent des signaux de danger qui activent les cellules immunitaires de la moelle épinière : la microglie. Celle-ci bascule alors en mode inflammatoire (M1) et produit un torrent de cytokines agressives, notamment le TNF-α et l’IL-1β (voir l'article : La microglie : Les gardiens immunitaires du cerveau au cœur de la neuro-inflammation)
Le β-caryophyllène du Copaïba possède ici une cohérence remarquable. Cette molécule agit sur les récepteurs CB2 présents sur la microglie, avec pour conséquence une diminution de plusieurs voies inflammatoires majeures comme NF-κB et l’inflammasome NLRP3.
En pratique, cela peut contribuer à diminuer la production de TNF-α et d’IL-1β, limitant ainsi l’entretien de la boucle inflammatoire qui bloque les pompes EAAT2 chargées d’éliminer le glutamate.
Les turmérones aromatiques du Curcuma longa semblent agir dans la même direction. En modulant également NF-κB, elles participent à réduire l’emballement inflammatoire tout en soutenant les mécanismes cellulaires de nettoyage des protéines anormales qui encombrent le motoneurone.
4.3) Protéger les mitochondries : préserver le dernier carburant cellulaire
Sous l’effet du calcium, du glutamate et des ERO, les mitochondries du motoneurone finissent par capituler. Or, sans ATP, l’immense axone du neurone moteur ne peut plus être ravitaillé correctement.
Les composés aromatiques du Romarin à cinéole, notamment le 1,8-cinéole et l’acide carnosique, possèdent ici un intérêt particulier. Ils semblent capables d’activer la voie Nrf2, véritable interrupteur de défense antioxydante au cœur de la cellule.
En stimulant cette voie de protection, le neurone augmente sa production de glutathion et d’enzymes antioxydantes, ce qui peut aider à préserver les mitochondries et maintenir une production minimale d’énergie.
Autrement dit : il ne s’agit pas seulement de calmer l’inflammation, mais de tenter de maintenir le motoneurone en capacité de survivre encore un peu plus longtemps dans un environnement devenu hostile.
5) Exemple pratique de protocole intégratif du terrain
Pour accompagner un organisme confronté à la SLA, la stratégie de terrain doit être d'une rigueur absolue. Elle ne cherche pas à remplacer le suivi neurologique indispensable, mais à saturer le terrain de molécules capables de nourrir les motoneurones et de calmer l'incendie immunitaire. L'approche est double : ravitailler les cellules par la micronutrition ET freiner la surchauffe par l'aromathérapie.
5.1) L'accompagnement micronutritionnel (nourrir, protéger et stabiliser)
- Magnésium L-Thréonate (ou Bisglycinate) : C'est le verrou indispensable. Il aide à freiner la surstimulation des récepteurs NMDA par le glutamate, limitant ainsi l'inondation toxique de calcium dans le motoneurone.
- Palmitoyléthanolamide (PEA) : La PEA agit en synergie parfaite avec le Copaïba pour calmer l'activation microgliale de la moelle épinière et protéger les structures nerveuses.
- Coenzyme Q10 (Ubiquinol) & Vitamine B3 (Nicotinamide) : Ce duo est le carburant direct de la chaîne respiratoire des mitochondries. Il vise à relancer une production minimale d'ATP pour soutenir le transport le long de l'axone.
- Oméga-3 (Riche en DHA) : Fluidifie les membranes des neurones et fournir les briques nécessaires à la résolution de l'inflammation
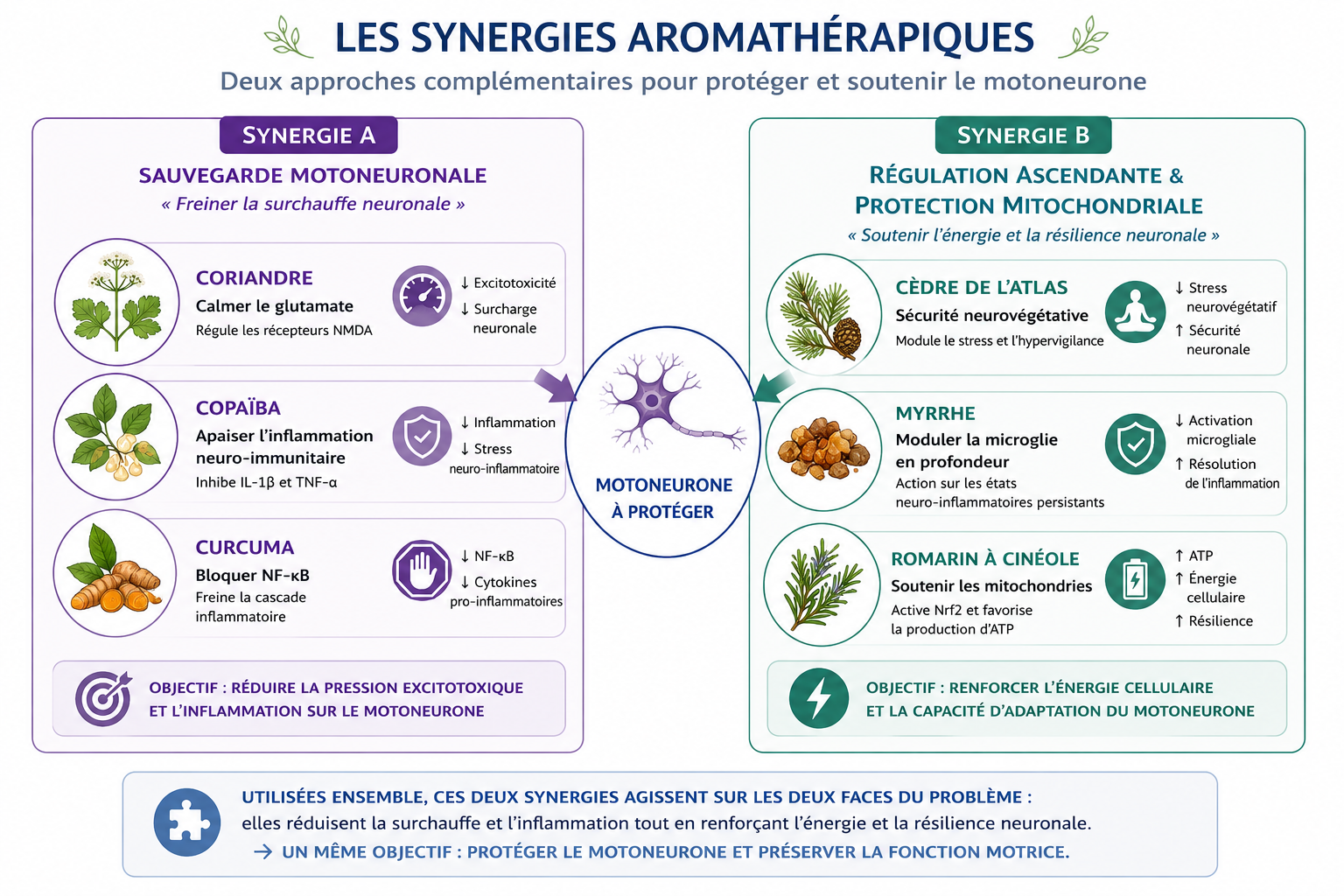
5.2) La synergie A : « Sauvegarde Motoneuronale »
Dans un flacon en verre ambré de 30 ml, assemblez la formule de haute régulation cellulaire suivante :
- HE Coriandre graines (Coriandrum sativum) : 30 gouttes (Bloqueur des récepteurs au glutamate)
- HE Copaïba (Copaifera officinalis) : 25 gouttes (diminution de production d'IL-1β et de TNF-α via les récepteurs CB2)
- HE Curcuma (Curcuma longa) : 15 gouttes (Inhibiteur de NF-κB, stoppant la fabrication des cytokines)
- Huile végétale de Jojoba ou de Macadamia : Compléter le flacon jusqu'à 30 ml
Protocole d’application :
- La voie cutanée (cible spinale) : Appliquer 6 gouttes du mélange en massage doux le long de la colonne vertébrale (du bas du dos jusqu'à la nuque) ainsi que sur la zone sous-occipitale (la base du crâne), matin et soir. Les motoneurones s'étendant tout au long de la moelle épinière, cette application locale permet une excellente diffusion des molécules volatiles vers les tissus nerveux cibles.
- La voie olfactive (apaisement central) : Pratiquer une respiration profonde directement au-dessus du flacon ou sur les poignets, 3 fois par jour, pour envoyer un signal de relaxation neurovégétative immédiat au tronc cérébral.
5.3) La Synergie B : « Régulation Ascendante & Protection Mitochondriale »
Dans un flacon en verre ambré de 30 ml, assemblez la formule ce second triptyque moléculaire :
- HE Cèdre de l'Atlas (Cedrus atlantica) : 25 gouttes
- HE Myrrhe (Commiphora myrrha) : 20 gouttes
- HE Romarin à cinéole (Rosmarinus officinalis ct cinéole) : 15 gouttes
- Huile végétale de Jojoba ou de Macadamia : Compléter jusqu'à 30 ml
Protocole d’application :
- La voie cutanée (cible spinale et nerveuse) : Appliquer 4 à 6 gouttes du mélange en massage doux le long de la colonne vertébrale (de la zone lombaire jusqu'à la nuque) ainsi que sur la zone sous-occipitale (la base du crâne), matin et soir.
- La voie olfactive (régulation du système nerveux autonome) : Pratiquer 3 grandes respirations lentes et profondes directement au-dessus du flacon ou en appliquant 1 goutte sur l'intérieur des poignets, 3 fois par jour.
Le Cèdre de l'Atlas (α-cédrol) : Le modérateur indirect du glutamate
L’α-cédrol (accompagné ici des atlantones, des sesquiterpènes apaisants) va agir « par le haut ». En provoquant un puissant switch neurovégétatif vers la dominance parasympathique, il envoie un signal de sécurité absolue au tronc cérébral et à la moelle épinière. Cette chute du stress sympathique diminue la réactivité de la microglie M1. Moins agressée, la microglie cesse de bloquer les astrocytes éboueurs, qui se remettent à aspirer le glutamate. C'est une baisse indirecte, mais globale, de l'excitotoxicité.
La Myrrhe (Furanosesquiterpènes) : Le bouclier lourd anti-IL-1β et anti-TNF-α
La Myrrhe contient des molécules uniques, les furanosesquiterpènes. Les études en neuro-inflammation montrent que ces composés inhibent puissamment la production de TNF-α et bloquent l'inflammasome NLRP3, le responsable de la maturation de l'IL-1β. De plus, la Myrrhe a une action protectrice directe contre la mort cellulaire (apoptose) des neurones hautement sollicités.
Le Romarin à cinéole (1,8-cinéole) : Le sauveteur de la mitochondrie via Nrf2
Le 1,8-cinéole franchit facilement les barrières biologiques. Dans le cas présent, le but est de stimuler la voie génétique Nrf2 au cœur du motoneurone. En activant cet interrupteur, il pousse la cellule à produire ses propres antioxydants pour protéger la cardiolipine des mitochondries. C'est l'outil indispensable pour maintenir la production d'ATP et soutenir le flux le long de l'axone.
5.4) Le protocole d'alternance clinique
Deux rythmes d'alternance sont particulièrement pertinents :
- Le rythme hebdomadaire (Le plus simple) :
- Semaine 1 : Synergie A (Coriandre / Copaïba / Curcuma) matin et soir.
- Semaine 2 : Synergie B (Cèdre / Myrrhe / Romarin) matin et soir.
- Avantage : Cela permet une imprégnation tissulaire stable de chaque totum, suivie d'une fenêtre de "redécouverte" cellulaire la semaine suivante.
- Le rythme des 3 jours (Le plus dynamique) :
- 3 jours avec la Synergie A, puis 3 jours avec la Synergie B, et ainsi de suite.
- Avantage : Idéal pour les terrains très réactifs ou en phase de forte fluctuation des symptômes, car cela empêche instantanément toute adaptation des récepteurs microgliaux.
L'application reste inchangée : 4 à 6 gouttes en massage doux sur la colonne vertébrale et la zone sous-occipitale, complétées par 3 grandes respirations au poignet.
Note importante : Les dosages et protocoles présentés dans cet article sont donnés à titre informatif pour un profil adulte type. Ils dépendent d'une évaluation globale de terrain et doivent impérativement être validés et ajustés avec votre neurologue, votre médecin traitant ou un praticien qualifié en aromathérapie clinique. Cet accompagnement de terrain ne se substitue en aucun cas à votre prise en charge médicale conventionnelle et nécessite de vérifier l'absence de contre-indications individuelles ou d'interactions médicamenteuses.
Conclusion : Offrir un sursis métabolique au vivant
La Sclérose Latérale Amyotrophique nous pousse à redéfinir notre regard sur les maladies neurodégénératives.
Limiter la maladie de Charcot à une fatalité électrique inexplicable ou à un simple « bug » génétique est une vision insuffisante. La SLA est l'illustration dramatique d'un écosystème cellulaire d'élite qui s'effondre sous le poids d'une surcharge métabolique et immunitaire.
Lorsque le motoneurone est attaqué par l'excès de glutamate, que ses mitochondries capitulent en mode de survie (CDR) et que la microglie de la moelle épinière devient pyromane en déversant ses cytokines, la cellule n'a plus les forces nécessaires pour entretenir son immense axone d'un mètre.
Il n'existe pas aujourd'hui de baguette magique pour effacer la maladie.
En revanche, la micronutrition et la biochimie végétale nous offrent des leviers d'une immense élégance pour modifier l'environnement de ces cellules en détresse.
En choyant la barrière intestinale pour stopper les fuites de LPS, en apportant les verrous nutritionnels contre le calcium, et en jonglant habilement avec la précision de nos synergies aromatiques — qu’il s’agisse de cibler frontalement les cytokines (IL-1β et TNF-α) ou de réveiller la sécurité neurovégétative et le secours mitochondrial —, on peut tenter de restaurer une homéostasie.
En rompant l’accoutumance des récepteurs par une alternance méthodique des molécules, on ne laisse aucun répit à l'inflammation et on offre à nos motoneurones le plus beau, le plus logique et le plus digne des sursis cellulaires.
© Guy Berlin - Aromatologue